全球暖化、氣候異常、極端天氣、海平面上升及海洋酸化等環境變遷現象,是二十一世紀影響人類社會永續與地球生態穩定的重要因素,也都與地球的碳循環息息相關。自工業革命以來,人類活動大量燃燒石油與煤炭,將數百萬乃至數億年前植物與藻類光合作用的產物,轉變成二氧化碳等溫室氣體排放到大氣中。才短短兩百年,在地球歷史上連一眨眼的時間都不到,大氣中二氧化碳濃度已經從工業革命前的280 ppm(每百萬分子)上升了50%到今日的420 ppm,相當於現在地球大氣總共有870拍克的碳(1拍克碳PgC=1015克碳 =10億公噸碳)。而且根據聯合國跨政府氣候變遷委員會(IPCC)的最新報告,2010–2019年人類活動的二氧化碳排放每年超過10拍克碳,儘管有全球陸域及海洋系統幫忙多吸收,最終大氣每年仍淨增加5拍克碳。
除了上述這些有機碳與無機的氣態二氧化碳,碳也以其它型式存在於不同的儲藏庫中,包含顆粒性的固態碳酸鈣(CaCO3)。在動物界,珊瑚的骨骼、許多軟體動物的殼與鳥蛋殼都是以碳酸鈣為主要成份。桃園沿岸的藻礁則是多細胞紅藻(無節珊瑚藻)鈣化所形成。然而地球上最大宗的碳酸鈣來源並非這些肉眼可見的鈣化生物結構,而是大洋中的單細胞微生物,包含異營性的有孔蟲與會行光合作用的鈣板藻。其中又以後者產量最多,每年達1.5拍克的碳被鈣板藻捕捉進碳酸鈣中,等於目前台灣年均碳排0.075拍克的二十倍。
鈣板藻顧名思義是會形成碳酸鈣板的藻類,彷彿每顆細胞住在自己的鈣板屋中,而且不同物種的形態各異(圖一)。雖然鈣板在細胞表面,但相較於其他生物鈣化主要是在細胞外形成結晶,鈣板藻則是在細胞內製造鈣板。鈣板藻在高基氏體產生的特殊囊泡中,打下橢圓的有機質基板,再利用帶負電的多醣類,調控鈣離子在基板邊緣形成一個個特定形狀的碳酸鈣結晶,個別的結晶排成一圈便是一片完整的鈣板,最後透過胞吐作用將鈣板送到細胞表面,一片一片砌成這顆細胞的鈣板屋。這種獨特的生物礦化過程,將本應長成菱面體的方解石,雕塑成小則1微米、大可至20微米的鈣板,還能運送到細胞膜外面,可說是自然界的一大驚奇。
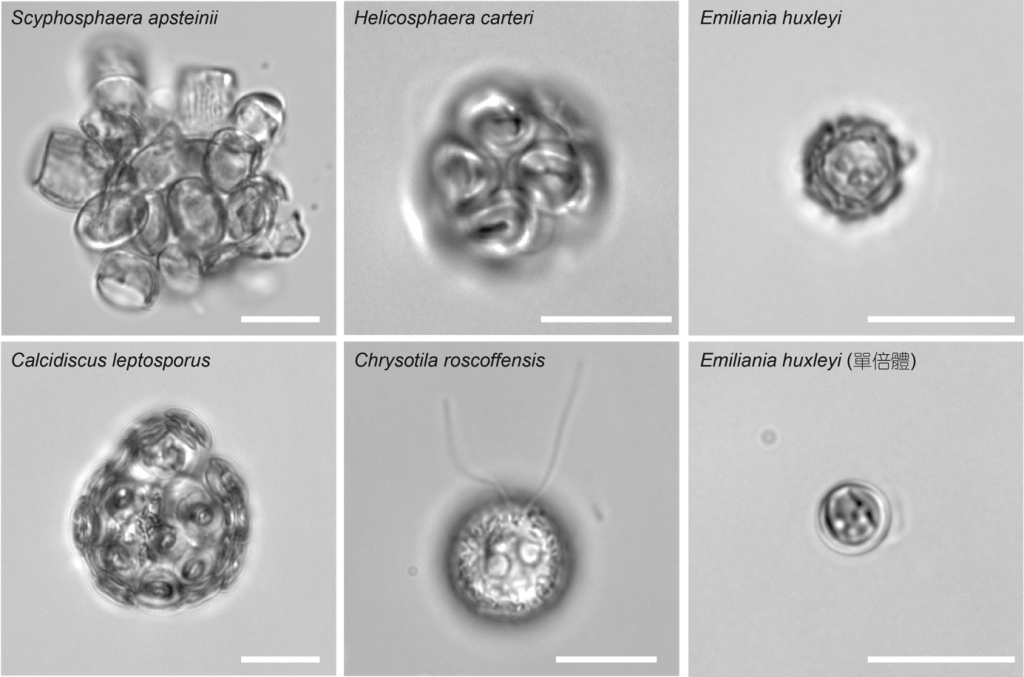
圖一:形形色色的鈣板藻,比例尺10微米。(王姿皓攝)
除此之外,鈣板藻最引人注目的,莫過於面積可達數十萬平方公里的藻華,由細胞最小但最常見的赫胥黎氏艾密利鈣板藻(Emiliania huxleyi,圖一)所形成。每年春夏之際在中高緯度海洋,艾密利鈣板藻獲得足夠營養鹽大量生長,細胞密度可達每毫升十萬顆,加上脫落鈣板的光線反射,在衛星影像中可以看到廣闊的海域被染成抹茶牛奶色。在全世界的艾密利藻華中,又以每年南緯38到60度夏季時形成的巨大方解石環帶最為壯觀,彷彿為南極洲裹上了一層厚棉被。藉著繁殖力強大的艾密利鈣板藻以及其它廣佈於各大洋的物種,鈣板藻的光合作用貢獻了海洋淨初級生產力的20%,相當於全球光合作用固碳(約100拍克)的10%。
鈣板藻死亡後仍可以持續影響碳循環。相較於微生物細胞或死亡碎屑所構成的顆粒性有機碳,碳酸鈣構成的顆粒性無機碳不會被異營性細菌使用分解,只要不是遇到高壓(例如4000公尺以上的深海)或酸性環境就不易溶解。換言之,顆粒性無機碳比顆粒性有機碳更容易從藻類生長的表水沉降到海底或深海,所以可以把從大氣及表水中固定下來的碳長期存放到其它儲藏庫。如果形成沉積物,鈣板將變成微米級的超微化石,能作為不同地質時期的指標化石,並紀錄下古海洋環境的故事。中生代晚期的白堊紀,正是因為當時鈣板藻大量產生的鈣板沉積經成岩作用變成白堊(chalk,以前粉筆的原料)而得名,而多年後這些白堊露出地表便成為風光明媚的景色,如英格蘭東南方的多佛白崖。此外,方解石密度(2.71 g/cm3)遠比海水(1.025 g/cm3)及細胞本身(~1.1 g/cm3)大,鈣板沉降時不只自己沉,連帶附著的有機物也會沉降得比較快、更容易到深海。這個壓艙石效應(ballast effect),不只是鈣板藻本身的細胞,其他藻類衍生的顆粒性有機物也可以一起搭便車,可說是生物固碳幫浦的加速器。
鈣板藻生物學還有其它與碳循環相關的秘密,是我們研究室關注的課題。首先,鈣板藻是一群具有單雙倍體世代交替的生物。人類等動物的生活史是雙倍型,單倍體細胞只存在於有性生殖的配子(精子與卵子)。鈣板藻與植物都有雙倍體及單倍體世代,但在植物中這兩者大多互相依存,而鈣板藻的雙倍體及單倍體則可以各自獨立,都能進行有絲分裂以無性繁殖。不同於一些單雙倍體外型類似的藻類(如:石蓴),鈣板藻的雙倍與單倍細胞的形態有顯著的二型性,尤其是在鈣板特徵方面,雙倍體通常有較大且結晶輻射狀的鈣板,單倍體則有較小且結構簡單的鈣板。不過最常見的艾密利鈣板藻是個例外,它的單倍體是有鞭毛但沒有鈣板的裸體細胞(圖一)。針對鈣板藻單雙倍世代交替的過程,我們研究室正利用基因體與其他體學方法,試圖找出單雙倍體世代表現不同形態與生理的關鍵。此外,在自然環境中不會鈣化的單倍體細胞有多常見,以及對艾密利鈣板藻整體鈣板產量有何影響,也是我們有興趣的研究方向。
近三十年來,科學家逐漸認識到海洋環境中的病毒,是促進微生物死亡與碳循環的重要因素。艾密利鈣板藻也有一個專門感染它的雙股DNA病毒,屬於俗稱巨病毒的核胞病毒門,基因體比一般感染人類的病毒大上許多,是SARS-CoV-2的14倍(相對的艾密利鈣板藻本身的基因體則只有人類的1/20不到)。在藻華後期的海水中,鈣板藻病毒爆發並導致大量宿主細胞死亡,故能增加有機與無機碳垂直向下傳遞的速率。不像SARS-CoV-2感染人類有輕重症之分,實驗室中一株艾密利鈣板藻一旦被無法抵抗的病毒株感染,整個族群最快數天就會完全死亡。為了解哪些遺傳因子決定它們之間的專一性,以及鈣板藻病毒是如何適應宿主,我們可以利用比較基因體學的方法來分析不同的宿主與病毒株。除了世代交替與病毒感染,我們研究室也透過篩選鈣化相關變異株與跨物種分析,試圖找出鈣化相關基因並進一步了解它們在鈣板藻的起源、演化與調控,不僅是為了認識它們的歷史,也有助預測全球環境變遷下鈣板藻的未來。
相較肉眼可見的生物,單一細胞的鈣板世界雖然微小,全球累積起來的數量與對生態的貢獻卻是舉足輕重。從南極海到北極海,從太平洋到大西洋,你可曾想過當你在呼吸排碳時,鈣板藻這兩億年來都勤奮地在固碳嗎?所以說下次去海邊或瞭望大洋時,可別忘了這些地球碳循環中的隱藏角色是多麽重要喔。
延伸閱讀
Balch WM. 2018. The ecology, biogeochemistry, and optical properties of coccolithophores. Ann Rev Mar Sci 10:71–98.
Frada MJ, Bendif EM, Keuter S et al. 2019. The private life of coccolithophores. Perspect Phycol 6:11–30.
Ku C, Sheyn U, Sebé-Pedrós A et al. 2020. A single-cell view on alga-virus interactions reveals sequential transcriptional programs and infection states. Sci Adv 6:eaba4137.
Laber CP et al. 2018. Coccolithovirus facilitation of carbon export in the North Atlantic. Nat. Microbiol. 3:537–547.
Liu YW, Eagle RA, Aciego SM et al. 2018. A coastal coccolithophore maintains pH homeostasis and switches carbon sources in response to ocean acidification. Nat Commun 9:2857.
Taylor AR, Brownlee C, Wheeler G. 2017. Coccolithophore cell biology: chalking up progress. Ann Rev Mar Sci 9:283–310.
IPCC. 2021. AR6 Climate Change 2021: The Physical Science Basis. https://www.ipcc.ch/report/ar6/wg1/
The Great Calcite Belt (Woods Hole Oceanography Institution). https://www.whoi.edu/multimedia/great-calcite-belt/
楊天南, 魏國彥. 2018. 鈣板藻與鈣質超微化石. 臺灣博物季刊 37:6–11.