香蕉是全球最重要的農業經濟產物之一,根據聯合國糧食及農業組織(Food and Agriculture Organization of the United Nations)的統計,2019年香蕉國際貿易達到135億美金的產值,香蕉產業養活了低度開發及開發中國家龐大人口。事實上,香蕉產業曾經面臨過全面性的崩潰,香蕉的瘟疫從20世紀中葉至今仍在蔓延,且有越來越嚴重的趨勢,所以過去十多年來曾有過未來將無香蕉可食用的預言。在1950年前人們食用的香蕉是一種稱為大麥克的品種(Gros Michel,美國人常稱為Big Mike),此品種香蕉深受美國人喜愛,是1950年代前進口美國的主要香蕉栽培品種。大麥克香蕉又大又甜且容易進行催熟處理,市場上戲稱大麥克香蕉等同於冰淇淋界的哈根達斯(Hägen-Dazs),而我們現在食用的香蕉則是夜市販賣的廉價冰淇淋。然而上世紀爆發的古巴專化型香蕉鐮刀菌黃葉病(Fusarium oxysporum f.sp. cubense或稱香蕉枯萎病、香蕉巴拿馬病),導致大麥克品種絕跡於全球市場;我們現在食用的香蕉是一種較能對抗鐮刀菌的品種,稱為卡文迪許(Cavendish)香蕉。
卡文迪許香蕉名稱的由來有個有趣的故事。在失去大麥克香蕉後,人們不斷地在世界各地找尋抗病又美味的替代品種,終於在英國威廉卡文迪許(William Cavendish)公爵的溫室找到一種三倍體的尖蕉(Musa acuminata)品種,據稱此品種是在1829年從模里西斯運往英國,幾經輾轉後由卡文迪許公爵購入收藏,沒想到在百年後竟成為拯救國際香蕉市場的主角。但是卡文迪許香蕉仍無法擺脫鐮刀菌的侵害,因為鐮刀菌不斷地突變,現今全球主要的感染源為第四型突變株(TR4;熱帶黃葉病4號;它在1967年首次出現於臺灣,隨後傳遍世界)。鐮刀菌的潛伏期可長達一年,致病孢子躲藏在土壤內,防堵困難。因此人類、香蕉、鐮刀菌三者之間的拉鋸戰仍持續進行中。育種專家希望能從野地發現新的抗病香蕉,或是利用人工雜交及生物技術等方法,培育出抗病品種。
香蕉泛指芭蕉屬(Musa)植物,與象腿蕉屬(Ensete)及地湧金蓮屬(Musella)組成芭蕉科(Musaceae),是單子葉植物中的一小分類群,所以香蕉是「草」(沒有木質化的樹幹)而不是「樹」。芭蕉屬由約70多種野生香蕉組成,它們具觀賞、工藝、及糧食等價值(圖一)。野生香蕉果實內含種子(圖一),雜交頻繁,育種專家將其歸類為四種常見的基因型,分別為A型來自野生的尖蕉、B型來自拔蕉(Musa balbisiana)、T型來自馬尼拉麻蕉(Musa textilis)、以及S型來自裂果蕉(Musa schizocarpa;種名源自它的果皮在成熟時開裂暴露出種子)。除了這四型外,種間雜交亦可發生在其他種類香蕉,例如:臺灣知名的香蕉育種專家邱輝龍博士以臺灣芭蕉(Musa itinerans var. formosana)為母本,拔蕉為父本,成功培育出新雜交品種,命名為Musa× formobisiana[1]。香蕉的種間雜交特性,以及頻繁且大量的育種工程,造就出許多雜交品種問世,但也讓育種史的追溯工作,具有一定的難度。

圖一、芭蕉科成員。(A)象腿蕉,圓柱形的假莖如同象腿般的筆直粗壯,拍攝於臺北植物園;(B)地涌金蓮,著名的佛教文化植物(感謝王瑞江博士提供照片);(C)紫夢幻蕉(Musa ornata),東南亞普遍的觀賞花卉(感謝葉育石提供照片);(D)馬尼拉麻蕉,其葉柄可提取纖維製造纜繩及造紙,拍攝於臺中農改場;(E)北蕉(Pei Chiao),尖蕉的栽培種之一,黃葉病高抗性,受蕉農喜愛,拍攝於屏東長治鄉;(F)無果籽的栽培種香蕉;(G)野生種香蕉的果實內含種實。
所幸香蕉擁有細胞質DNA異源特性。在植物細胞內,我們稱細胞核以外的物質為細胞質,而細胞質內含兩種胞器: 粒線體及由質體(Plastid)所特化的葉綠體(圖二)。在細胞內除了細胞核攜帶DNA遺傳物質外,粒線體和葉綠體也持有自己的基因體,我們將粒線體及葉綠體的DNA稱為細胞質DNA。在自然界中,多數陸生植物的細胞質DNA為母系遺傳,也就是它們的粒線體及葉綠體皆遺傳自母親,所以稱為細胞質DNA同源特性;但是有極少部分的植物,例如:某些針葉樹、黃瓜、甜瓜,其細胞質DNA為異源性,也就是它們的粒線體及葉綠體遺傳自不同的親本。香蕉的細胞質DNA的異源特性在1994年被證實[2],其粒線體遺傳自父親,葉綠體則來自母親,此種異源遺傳現象提供育種專家追溯香蕉雜交史的研究途徑。此後,大量且複雜的專一性PCR引子被開發且應用在香蕉育種研究。
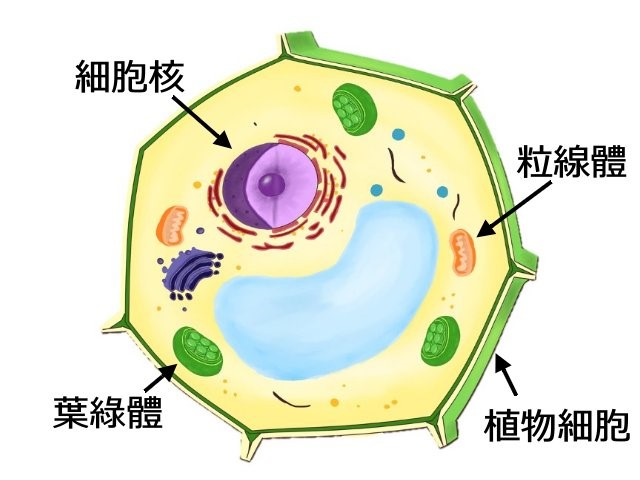
圖二、植物細胞構造圖(感謝曾韵婷小姐繪圖)。
在後基因體時代(Postgenomic era),人們已無法滿足少數DNA片段所提供的遺傳訊息,隨著次世代定序技術的進展,以全基因體資料進行研究成為共識。全基因體不僅提供完整且不偏頗的資料,其豐富的遺傳訊息亦可強化統計運算的樣本可靠性。然而生物的核基因體多樣且複雜,組裝核基因體序列與後續的分析需要龐大的電腦運算及人工判讀,且食用香蕉多為多倍體品種,增加核基因體組裝的困難度。因此,我們研究室把目標鎖定在粒線體及葉綠體上。如上所述,此兩種胞器持有自己的基因體,且它們的基因體具有多拷貝特性(也就是:一個胞器內有相同的幾份基因體)。植物胞器基因體拷貝數在不同的發育階段及不同組織部位,呈現極大的差異。一般而言,發育中的組織含有豐富的胞器DNA,而成熟部位則含量稀少。提高目標DNA的含量有利後續的組裝及提昇資料的可信度。
我們從剛發育的香蕉幼芽抽取DNA,進行基因體淺層定序(Genome skimming),也就是說我們只定序少量的資料,並利用幼芽組織含有大量胞器及胞器基因體多拷貝的特性,使粒線體及葉綠體DNA的定序資料在總資料上佔有相對較高的含量,且此資料含量足夠組裝出高可信度的粒線體及葉綠體基因體序列。基因體淺層定序有幾項優點:(1)在有限的研究經費下,不需大量定序資料即可取得豐富的基因體遺傳訊息;(2)節省單一物種的定序經費,因此研究的範疇可涵蓋多物種,例如: 研究計畫可取樣整個分類屬(Genus)底下的物種,充分評估該分類層級的基因體演化多樣性;(3)定序資料相對較小,不需要繁重的電腦運算及人工判讀。以香蕉為例,我們只需約0.8Gb的定序量,以4核心筆電運算,約2小時可獲得定序深度超過50倍及1,000倍的粒線體及葉綠體基因體組裝,這些組裝雖可能不完整,但足夠提供百分之百的基因體編碼序列/區域訊息,將這些編碼區域串聯成資料矩陣則可進行演化分析。
我們以粒線體及葉綠體基因體的編碼區域建構親緣演化樹(圖三),結果顯示象腿蕉屬與地湧金蓮屬為姊妹群關係,與芭蕉屬關係較疏遠。芭蕉屬內可再細分成兩組分支群(Clade),雜交育種常使用的尖蕉與拔蕉被歸類在分支群I,而常被誤認為拔蕉的馬尼拉麻蕉則被劃分在分支群II。令人驚訝的是,粒線體演化樹表明臺灣芭蕉與拔蕉的雜交品種Musa × formobisiana與拔蕉的親緣關係最為相近,但葉綠體演化樹卻支持此雜交品種是臺灣芭蕉的最近似親緣物種。這個不一致的結果反映出香蕉細胞質DNA的異源特性(父系遺傳的粒線體及母系遺傳的葉綠體),也符合Musa × formobisiana的育種史。以此方法我們亦可推論千層蕉(Musa × chiliocarpa)是以尖蕉為父本,拔蕉為母本的雜交品種。
在香蕉的親緣分析上,我們發現粒線體基因體明顯比葉綠體基因體具備更為準確的解析度。粒線體演化樹揭示尖蕉及其衍生的栽培種組成一單系群(Monophyletic group)─亦即它們只有一個共同的祖先。但葉綠體演化樹卻全盤混淆了它們與其他種香蕉的關係(圖三)。尖蕉的栽培種多在近百年內被建立,母系遺傳的葉綠體在短時間內累積的DNA突變不足以區分尖蕉與其他種香蕉,而父系遺傳具有快速累積DNA突變的特性,我們的研究成果揭露父系遺傳的粒線體基因體是研究香蕉親緣關係的利器。未來若能收集芭蕉屬內所有70多種香蕉的DNA,以基因體淺層定序組裝粒線體基因體,有效釐清各種香蕉的親緣關係,將可加速香蕉雜交育種工程的進展。
大麥克香蕉的事件體現了維持物種遺傳多樣性的重要。人們為了追求高效益,有目標性地窄化香蕉的遺傳多樣性,只栽種單一品種/系香蕉,一旦發生瘟疫,整個產業將面臨崩潰的命運。為了讓後代子孫可享用甜美的香蕉,策略性的選擇各地品種雜交育種是擴大香蕉遺傳多樣性的可行方法。政府與企業應鼓勵蕉農經營非單一品種栽植,提供市場多樣性的香蕉。而我們研究團隊的研究成果可作為追溯香蕉雜交育種史的重要參考依據[3]。
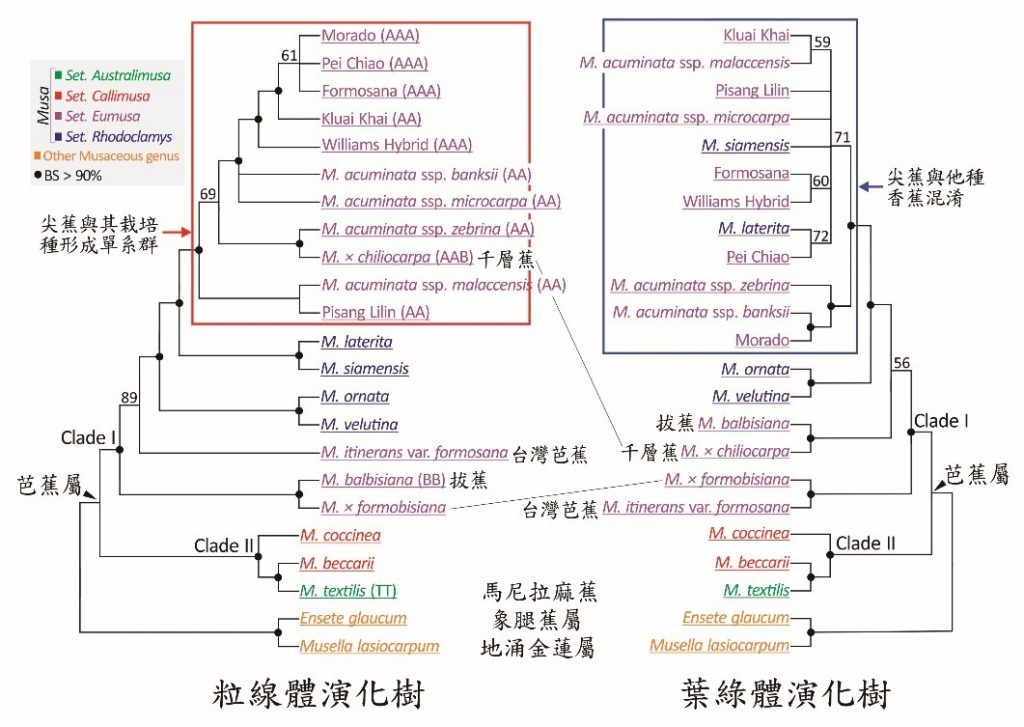
圖三、香蕉的親緣演化分析。
致謝
感謝屏東香蕉研究所及行政院農業委員會農業試驗所提供香蕉品種樣本,本院生物多樣性研究中心和中研院主題研究計畫經費支持。
延伸閱讀
[1]Chiu HL, Shii CT, Aleck Yang TY. Taiwania 2017, 62, 147–150. Doi: 10.6165/tai.2017.62.147.
[2]Fauré S, Noyer JL, Carreel F, Horry JP, Bakry F, Lanaud C. Curr. Genet. 1994, 25, 265–269. Doi: 10.1007/BF00357172.
[3]Wu CS, Sudianto E, Chiu HL, Chao CP, Chaw SM. Front. Plant Sci. 2021. Doi: 10.3389/fpls.2021.713216.