碳化稻米揭露臺灣五千年前農耕文化
稻米,是臺灣現今食文化中最重要的主食,而水稻亦為臺灣當前最主要的糧食作物;若以分類學來著眼,亞洲栽培稻(Oryza sativa)大致可區分為秈稻(indica) 與稉稻(japonica)兩種亞型。
臺灣近代稻作栽培深受日本文化影響,以稉稻為主要栽培稻,但其實臺灣稻作栽培文化悠久歷史。根據南科考古研究的結果,遺址分布於現今臺灣南部科學工業園區臺南園區,而南關里、南關里東遺址所代表的大坌坑文化,根據碳14年代偵測結果顯示,可追溯至距今約5000年前左右。考古遺跡中,發現成千上萬臺灣最早的稻米及小米碳化種子,並根據種子大小及外觀,推測臺灣在新石器時代已具有選種的技術;出土文物中也發現石刀、石鐮、貝刀等常被視為農耕工具的史前器具。這些證據盡揭示了5000年前臺灣已經進入農業時代,藉由研究這些化石及文物,再結合語言學及稻米適應(adaptation)基因的研究,可以讓我們更瞭解南島語族的遷徙過程與文化關係(1)。
臺灣栽培稻進程與山地原住民陸稻品系
稻是臺灣住民的主食。臺灣的栽培稻作歷史可分為三個時期:一、臺灣原住民所栽種的山地陸稻,這些陸稻品系與南島語族遷徙有關;二、明末清初時期,由於大量漢人移居臺灣從事開墾,自中國沿海地區引進許多秈稻品系(亦稱為在來稻);三、近代臺灣主要習慣食用稉稻(亦稱蓬萊稻),主要受日本稻作文化影響,自西元1895~1945年日治時期,由日本育種專家如磯永吉博士等引種、改良溫帶型稉稻,以適應臺灣環境及氣候(2)。
十九世紀初,由日本育種學者們至臺灣高山原住民部落,蒐集近60種山地陸稻。可惜這些珍貴的山地陸稻品系由於年代久遠,歷史背景資料大多亡佚。經語言學家本院李壬癸院士、法國國家科學院Sarget博士等藉由語言辨識品系名稱,目前已鑑別出十多種搜集品系屬於何種臺灣原住民族;而後,各地育種學者如葉茂森博士、劉啟東博士、羅正宗博士、宣大平博士等在臺灣中部、南部以及花東地區等地蒐集各地方陸稻品系,迄今已累積超過百種山地陸稻品系。這些山地陸稻品系外表型差異顯著,包括植株之株型、高度、葉片之顏色及寬度、分蘗數及分蘗角度、莖桿粗細程度、穗長、抽穗期長短、倒伏程度、芒的有無及長短、落粒性強弱、穀粒形狀及大小、穀粒顏色、種皮顏色等都具有極大的差異(圖 1)。且這些山地陸稻品系中,許多具有抗病害、抗蟲害、耐旱、耐鹽、抗紫外線(UV)等適應自然環境的各類特性,未來可成為有助於改良栽培稻的重要遺傳資源。目前,這些山地陸稻品系已逐步完成全基因體定序,藉由親緣分析研究及稻適應相關基因的研究,可提供釐清臺灣原住民與水稻適應歷史、南島語族遷徙的重要佐證(3)。

圖 1. 各種山地陸稻外表型。
水稻的適應相關基因、過程與育種
稻米的適應歷史是十分複雜的過程,透過人類不斷地選拔育種,選擇喜好的外表型,如無芒的種子與低落粒性的品系,採收過程較為方便順利;選育理想株型包括分蘗數與分蘗角度,較直立株型可使栽植密度提高,而增加產量。20世紀的綠色革命,選育出半矮性品系IR8,針對植株倒伏問題進而增進產量的半矮性狀。另外,育種家也選育適合當地緯度、日光長度及溫度等環境條件的品系等。學著們可藉由追溯這些適應相關基因在各地方品系及栽培稻、野生稻之間的差異,輔以語言學、考古學、歷史文獻等,而建構出水稻適應的歷史。
短日照植物透過種子形式以度過冬季冷冽的環境,繁衍下一代;因此短日照植物感應日長時間變短後,即開花並結果。水稻為短日照植物,為適應不同緯度地區的日照長度,自然界中水稻抽穗期長短也存在差異;於短日照環境可正常抽穗,而長日照環境下則較慢抽穗。為適應栽培面積廣大的亞洲地區,其中最主要的因素為不同地區及不同季節的日長及溫度變化,進而被育成適應溫帶、熱帶不同環境氣候的多樣化水稻品系。其中,調控抽穗期的基因超過十個以上,且序列中存在許多變異,可能造成胺基酸的改變或影響基因表現,進而改變抽穗期長短。而抽穗期長短更是直接影響一年的收成次數,左右稻米產量。其中主要影響抽穗期長短的基因Heading date 1(Hd1)基因位於水稻第六條染色體:短日照環境下,Hd1促使下游基因表現,使植株提早開花;而長日照環境下,Hd1抑制下游基因表現,延遲植株開花。藉由此調控機制,育種家選育出適應各地區不同栽培方式以及不同收穫次數的水稻。
但當Hd1基因失去功能,導致水稻對於光週期變化不敏感,使抽穗期的調控不再受日長影響;例如位於亞熱帶地區臺灣而言,水稻收穫將因此由每年一次收成而增加到兩次收成。以水稻品系日本晴為參考序列,Hd1有兩個外顯子(exon),蛋白質共有395個胺基酸。經由比對山地陸稻品系以及數個現代栽培品系全基因體序列,再結合了來自國際稻米研究所(International Rice Research Institute, IRRI)所完成的約3000株水稻全基因體定序資料中,發現有些品系在第一個外顯子有15個單核苷酸多型性突變(Single Nucleotide Polymorphism, SNP)以及兩個插入突變(insertion);有些在第二個外顯子上有三個單核苷酸多型性突變;或有一個大片段1901鹼基對之反轉錄轉位子(retrotransposon)序列插入。上述多數的突變皆造成胺基酸的改變,尤其是發生於重要功能區域的突變,進而影響Hd1蛋白表現的差異甚至失去蛋白功能。我們分析收集到的材料中,約有十個山地陸稻品系在Hd1第二個外顯子上皆因為1901鹼基對的插入事件而導致完全失去功能。而此1901鹼基對序列另外在水稻基因體中第五及八條染色體上各有相似的序列,經比對資料庫後確實屬於反轉錄轉位子。
前人研究(4)中指出著名水稻品系臺中65號在Hd1基因相同位置處插入相同的1901鹼基對反轉錄轉位子。但臺中65號係由日籍育種學者末永仁博士於1924年以龜治與神力兩種日本水稻品系雜交育種而成,此兩種日本水稻品系皆不含此1901鹼基對插入突變且皆對於光週期敏感,卻育成帶有插入突變且對光週期不敏感之臺中65號,引起我們想更進一步研究的興趣。經由比對山地陸稻品系以及其他超過3000株水稻全基因體定序資料,此1901鹼基對插入突變僅在近十個的山地陸稻品系及臺灣的現代品系(或以臺灣品系為親本之品系)中發現。因此,我們藉由此全基因體定序分析結果可歸納出:日治時期至今,農業試驗單位肩負保存種原以及維持種原活力的任務,對於這些山地陸稻品系每隔十年將會復育種植而後保存種子;同時,末永仁博士期望在臺灣培育出符合日本人嗜好口感之日本型水稻,在當時的臺中州農事試驗場進行選育龜治與神力兩品系雜交育種成臺中65號。在相近試驗田間復育陸稻及選育臺中65號的過程中,發生基因滲入(introgression)而導致山地陸稻品系之基因體滲入臺中65號基因體中(5)(圖 2)。如此美麗的錯誤卻選育出具備高產量、高品質、抗稻熱病、適應力強且對光照不敏感之品系,在臺灣第一、二期作皆適合栽種,逐漸成為臺灣水稻栽培的主流。目前臺中65號被利用在臺灣近90%稉稻品系的譜系,影響深遠。
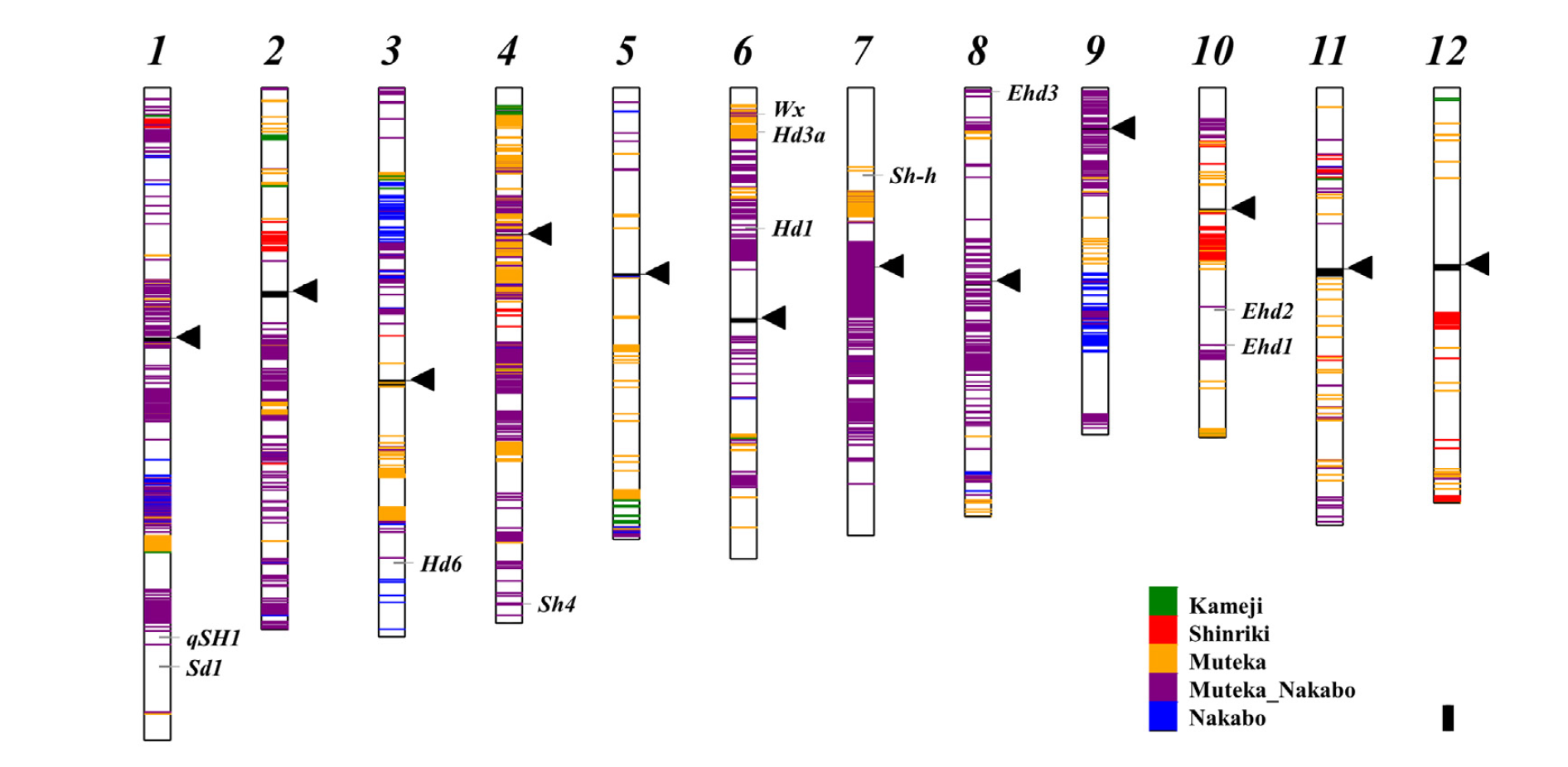
圖 2. 臺中65號基因體中,各山地陸稻品系特定基因體區域分布。在水稻的12條染色體中,綠色表示龜治基因體區域、紅色為神力、橘色為山地陸稻Muteka、藍色為山地陸稻Nakabo,而紫色為Muteka與Nakabo共有的區域。其中第六條染色體短臂中間即為Hd1基因。
隨著全球氣候暖化,農業用水比例逐漸降低,陸稻抗旱功能的品種特性也逐漸受到重視。目前臺灣陸稻主要栽培地區分布於臺東縣、花蓮縣、南投縣等山地原住民部落。以臺東農業改良場為例,於民國53年命名推廣以臺中65號為母本、東陸育11號為父本之東陸1號,具有抗稻熱病、不易倒伏、耐旱等特性,適合缺水地區栽培;其他如東陸2號、東陸3號等也都具有山地陸稻耐旱的特性(6)。雖然陸稻的產量只有一般水稻的30-40%,且米質不若良質水稻栽培品種,但其對於自然環境的抗性及耐受性高,適合做為水稻的育種材料,若能利用這些資源育出灌溉需求較低的水稻,在有限的水資源條件下,將獲得最大的稻作生產效益。
未來展望
未來的研究將更進一步了解適應基因於臺灣山地陸稻品系的基因體序列變異程度以及適應相關性狀差異,有助於揭示臺灣在地稻米品系的適應歷史。同時,比較臺灣稻米與鄰近國家包括日本、韓國、中國、中南半島以及南洋群島之地方稻米品系,將揭開南島語族遷徙與傳播作物的歷史淵源。此外,這些臺灣山地品系較現今栽培水稻生育所需用水較少,且抗病性、抗蟲性及生長勢皆優於現今栽培水稻,未來將利用作為基因庫來源,有助於選育出更符合環境需求之稻作。
參考資料
- J. S. Hsieh et al., Studies on Ancient Rice-Where Botanists, Agronomists, Archeologists, Linguists, and Ethnologists Meet. Rice 4, 178-183 (2011).
- 鄧耀宗, 臺灣稻作之回顧與展望. 高雄區農業改良場研究彙報 14, 1-23 (2003).
- L. Sagart et al., A northern Chinese origin of Austronesian agriculture: new evidence on traditional Formosan cereals. Rice (N Y) 11, 57 (2018).
- K. Doi et al., Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev 18, 926-936 (2004).
- F. J. Wei et al., Both Hd1 and Ehd1 are important for artificial selection of flowering time in cultivated rice. Plant Sci 242, 187-194 (2016).
- 丁文彥, 陸稻-東陸1,2,3號品種介紹. 臺東區農業專訊 79期, 8-11 (1992).